基因敲除原理和方法-基因敲除原理方法
 6人看过
6人看过
基因敲除原理与方法:现代生物学的基石与前沿
基因敲除(Gene Knockout, KO)是分子生物学、遗传学和发育生物学中最核心、最有力的实验手段之一。它经由人为地在基因组中制造基因缺失或功能丧失,从而直接验证基因在生物体中的功能,并解析复杂的生物性状与基因型之间的因果联系。从早期的随机突变筛选到如今的 CRISPR 系统,基因敲除技术的不断革新,极大地推动了我们对生命本质认知的深化。
基因敲除的基本原理
基因敲除逻辑在于"功能缺失即表型缺失"。当我们成功敲除一个基因时,该基因原本编码的蛋白质将不再被合成,或者其表达量的显著减少。在生化层面,这会导致依赖于该蛋白质的下游信号通路发生断裂;在细胞层面,导致细胞周期停滞、凋亡或功能障碍。
核心机制
结构破坏:通过手段切断基因序列,阻止转录或翻译。 调控缺失:破坏基因的启动子或增强子,使基因表达低下。 插入突变:在基因关键区域插入非编码序列,导致移码突变或提前终止。 同源重组:利用同源重组机制,将外源 DNA 替换为自身 DNA,从而造成基因缺失。基因敲除的主要方法
根据对基因序列的修饰方式和效率不同,基因敲除技术主要分为以下两大类:
传统方法:同源重组(Homologous Recombination, HR)
这是最早用于哺乳动物细胞及体细胞敲除的技术,依赖于细胞自身的同源重组修复机制。
原理:设计一段与目标基因上下游序列高度同源的 DNA 片段,将其整合到基因组中。在细胞分裂过程中,细胞会利用这段同源序列,以自身 DNA 为模板,置换出目标基因序列,导致基因缺失。
长处:操作简便,无需复杂的合成酶,可结合到小鼠基因组中特定的位点。
局限:效率较低(仅约 10%),且在小鼠胚胎干细胞中产生多个非同源重组位点,导致基因型复杂化。
| 数据类型 | 技术名称 | 优点 | 缺点 |
|---|---|---|---|
| 基因敲除小鼠 (KO) | 同源重组 (HR) | 操作成熟,成本低,可结合特异性标记 | 效率低,重组位点多,需大量动物 |
| 基因敲入 (KO) | 同源重组 (HR) | 可引入点突变,改变蛋白结构 | 效率低,操作复杂 |
| 基因敲除小鼠 (KO) | CRISPR-Cas9 | 效率高,效率优于 HR | 脱靶风险,需设计引导 RNA |
| 基因敲入 (KO) | CRISPR-Cas9 | 效率极高,灵活性强 | 脱靶风险,脱靶编辑需验证 |
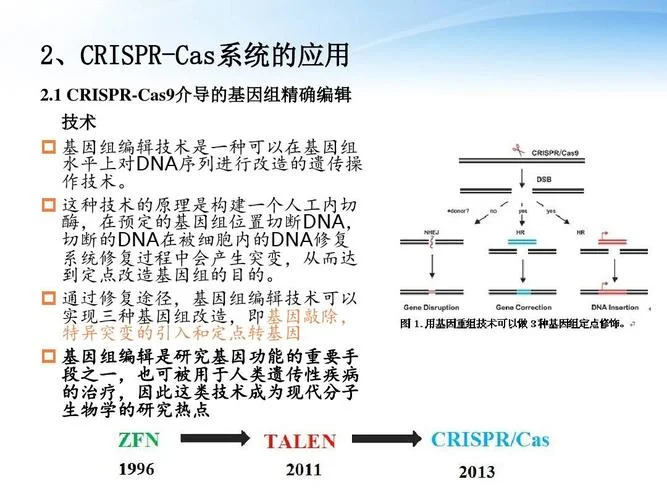
新型高效方法:CRISPR-Cas9 系统
近年来,CRISPR-Cas9 技术因其很高的精准度和效率,迅速取代了同源重组,成为首选的基因敲除工具。原理:利用 Cas9 核酸酶(向导 RNA 引导)识别基因组特定位点,造成双链断裂。细胞随后通过非同源末端连接(NHEJ)或同源修复(HDR)途径完成修复。
NHEJ 途径:导致插入或缺失(Indels),直接破坏阅读框,实现基因敲除(KO)。
HDR 途径:利用供体模板进行精确修复,实现基因敲入或精确修饰。
特长:
效率高:单次操作即可产生大量杂合或纯合敲除细胞系,效率可达 50% 以上,远超传统 HR。
灵活性:可定点设计,精确控制切割位点。
便捷性:无需构建复杂的筛选载体,通过转染即可获得细胞系。
挑战与对策:
脱靶效应:切割非目标基因位点。
对策:设计多序列的 gRNA 提高特异性;使用 PAM 序列识别机制;经过测序验证。
供体模板递送困难:HDR 效率低于 NHEJ。
对策:使用双底 gRNA 引导 NHEJ 和 HDR;优化递送系统(如 AAV 病毒);优化供体模板设计。
基因敲除技术的评估指标
为了确保敲除的准确性,生物学家会采用以下指标进行评估:
1. 杂合子率:在基因型为杂合子(+/KO)的细胞或小鼠群体中,观察到纯合敲除个体(+/+)的比例。
2. 过表达率:在基因型为杂合子(+/KO)的群体中,观察到纯合野生型细胞(+/+)的比例。
3. 存活率:基因型为纯合敲除个体(+/+)在特定培养条件下存活的比例。
4. 特异性:敲除后,目标基因的表达量是否显著降低,而非目标基因的表达是否受影响。
基因敲除技术作为研究生命奥秘的利器,已经从简单的随机断裂进化为一种高度精密、多功能的分子手术刀。从同源重组到 CRISPR-Cas9,每一次技术的迭代都拓展了我们探索生命的边界。随着 sgRNA 设计算法和递送系统,基因敲除技术将在基础研究和临床转化中发挥更为关键的作用,持续驱动医学和生物学。
 47 人看过
47 人看过
 44 人看过
44 人看过
 43 人看过
43 人看过
 32 人看过
32 人看过